

Ex-vivo LPS stimulation model coupled with quantitative PCR and its multispecies application in immunonutrition
Posted: 22 February 2010 | Dr Afif Abdel Nour, Associate Professor in Molecular Biotechnology and qPCR expert, Institut Polytechnique LaSalle Beauvais, France. Contributing Authors: Dr Flore Dépeint, Research Associate, specialised in Clinical and Preclinical Nutritional Studies and Gastrointestinal Health, Institut Polytechnique LaSalle Beauvais, France; Professor Elie Barbour, Professor of Veterinary Microbiology, Department of Animal and Veterinary Science, Faculty of Agricultural and Food Sciences (FAFS), American University of Beirut (AUB), Lebanon and Dr Philippe Pouillart, Associate Professor in Immunonutrition, Institut Polytechnique LaSalle Beauvais, France
Disorders of the immune systems leading to chronic inflammation and allergies are increasing in modern societies. While the possible causative factors are both environmental and nutritional, prevention and even curative options may be derived from our diet. Because background levels of cytokine expression in the general population are generally low, this model was developed to mimic an acute pro-inflammatory threat by a bacterial lipopolysaccharide (LPS).
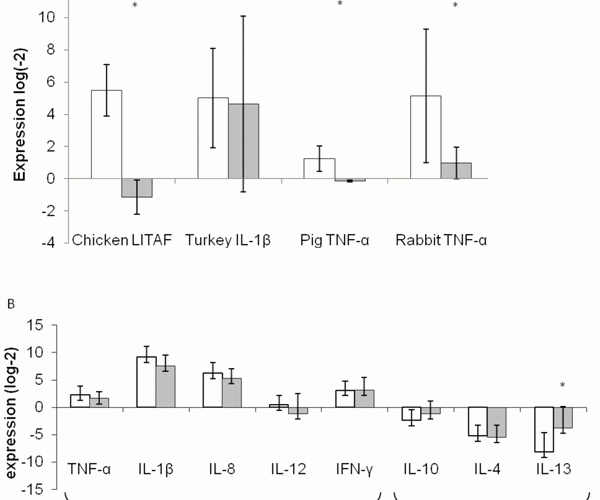
Figure 1 Cytokine expression: Relative expression of TNF-α or related cytokine in four different animal models fed a fiber-enriched diet or a dextrose placebo (Fig 1A, top). Relative expression of a panel of human cytokines following four weeks with a fiber-rich diet compared to a maltodextrin placebo (Fig 1B, bottom). Results are expressed as Mean±SEM. * represents statistically significant variation between the two groups (p< 0.05).
Disorders of the immune systems leading to chronic inflammation and allergies are increasing in modern societies. While the possible causative factors are both environmental and nutritional, prevention and even curative options may be derived from our diet. Because background levels of cytokine expression in the general population are generally low, this model was developed to mimic an acute pro-inflammatory threat by a bacterial lipopolysaccharide (LPS).
The development of a rapid and accurate method is critical for quantifying imbalances in the expression of multiple genes in clinical research. Quantitative real time PCR is used to measure variations in the expression of target genes involved in immunological pathways, normalised to an internal reference gene.
The model was tested in preclinical studies on four species: chicken, pig, turkey, and rabbit, as well as in a clinical trial. Subjects were fed fiber-enriched diets for a set length of time. At the end of the feeding period, freshly drawn whole blood was exposed to the antigenic factor for six to 24 hours, during which time the immune response is triggered. The fiber tested in preclinical studies reduced the inflammatory process (TNF-α, IL-1β), while the fiber tested in the clinical trial modulated both the Th1 and Th2-related pathways significantly.
We showed that this protocol was both rapid and accurate, and could be applied to monitor the effect in zootechnic as well as clinical interventions.
1. Introduction
Acute and chronic imbalance of the immune response is a growing problem in worldwide population, whether it is expressed as chronic inflammatory disorders or allergies. Changes in dietary habits in the recent decades may be a cause of those disorders1-4, it may include dietary changes such as an increased intake of refined sugars and fat, associated with reduced amount and variety of fiber as well as fruits and vegetables. While the most part of the immune programming is done during the early years of life, it is still likely that adopting appropriate dietary habits may help prevent or at least modulate those effects in later life, when disorders arise.
Several techniques are available to assess immunological responses. Direct measurement of circulating cytokines can be performed by ELISA tests on plasma samples. Except for pathological settings, where the immune response is increased, the background levels of circulating cytokines and other immune signaling molecules tend to be low, and significant variation are almost impossible to detect5. Animal models using LPS infusion leading to endotoxemia have been used successfully to study the mechanisms of acute inflammation and investigate potential treatments6. Ex-vivo LPS stimulation can also be applied, allowing to investigate a large number of hypotheses without the toxic risks to the subject. In this case leukocytes are generally isolated and cultured with LPS in a synthetic cell culture medium6. After stimulation, the cytokine production can be measured directly by ELISA protocols, through detection of the cytokines being released into the culture medium5. An alternate method is to measure variations in the genetic transcription using quantitative PCR after RNA extraction from the cellular pool.
We present in this article a method using whole blood stimulation and quantitative PCR (qPCR). This method was used to monitor the effect of dietary fibers on the expression of TNF-α or a closely related cytokine in five species: chicken (Gallus gallus), pig (Sus scrofa domestica), turkey (Meleagris gallopavo), rabbit (Oryctolagus cuniculus) and human (Homo sapiens). In addition, a wider panel of cytokine modulation was tested in the clinical study.

Figure 1 Cytokine expression: Relative expression of TNF-α or related cytokine in four different animal models fed a fiber-enriched diet or a dextrose placebo (Fig 1A, top). Relative expression of a panel of human cytokines following four weeks with a fiber-rich diet compared to a maltodextrin placebo (Fig 1B, bottom). Results are expressed as Mean±SEM. * represents statistically significant variation between the two groups (p< 0.05).
2. Material and methods
2.1 Experimental protocols
Preclinical and clinical trials were approved by local ethic committees and follow local and national guidelines. For the clinical trial, all subjects were exposed to the written informed consent of the protocol, and their approvals were secured.
Preclinical studies: Four separate studies were performed, each on a different animal species. A common element, however, was present in all setups, allowing for cross-species comparison. Indeed all animals were fed the same test ingredient, a dietary fiber derivative at a concentration of 30 ppm, from hereon called Fiber 1. Species and feeding protocols are detailed in Table 1.
Clinical trial: Healthy volunteers, aged 18 to 24, were given a dietary fiber supplement at a dose of four g/day, from hereon called Fiber 2, for a total duration of four weeks.
For both settings, a control group was introduced, where the fiber was replaced by an isocaloric amount of dextrose. The homogeneity of diets between the placebo and test groups was ensured in the preclinical trials by the use of a defined synthetic diet. In the clinical trial, homogeneity between the two groups was assessed through food intake questionnaires beforehand and regular food diaries during the intervention period.
| Species | Target gene | Age, length of feeding period |
| Chicken (Gallus gallus) | LITAF (Th1) | Birth, 105 days |
| Turkey (Meleagris gallopavo) | IL-1β(Th1) | Birth, 105 days |
| Pig (Sus scrofa domestica) | TNF-α (Th1) | Birth, 48 days |
| Rabbit (Oryctolagus cuniculus) | TNF-α (Th1) | 28 days, 42 days |
| Human (Homo sapiens) | TNF-α, IL-1β, (Th1) IL-10, IL-13 (Th2) | 18 -24 years, 28 days |
Table 1: Summary of experimental conditions for preclinical studies Animal species, selected cytokine target genes and basic information on the feeding protocols are presented here.
2.2 Ex-vivo LPS stimulation and quantitative PCR
At the end of the feeding period, blood samples were collected in heparin tubes. Aliquots of 1mL whole blood were incubated with 100 μL of 10 ng/mL lipopolysaccharide extracted from E. coli (Sigma, Lyon, France) at 37°C under permanent rotation (150 rpm).
After incubation, 0.5 mL blood was stabilised in 1.3 mL RNeasy protect cell solution (Qiagen Courtaboeuf, France) before extraction. RNA from chicken, rabbit and human samples were extracted using RiboPure Blood Kit (Applied Biosystem, Courtaboeuf, France), while those from pig and turkey were extracted using RNeasy Lipid Tissue Mini Kit (Qiagen) according to manufacturer’s instructions. All extracts (1 mg RNA) were then processed using Quantitech Reverse Transcriptase kit (Qiagen) according to manufacturer’s instructions for reverse-transcription. After each step, quality and concentration of the extracts (RNA, cDNA) were assessed using the spectrophotometer method with Helma UV cell (Eppendorf, Paris, France). Optical density (OD) at 260 nm and 280 nm were measured in triplicate to estimate RNA and cDNA concentration and quality.
The expression of cytokines of interest were analysed and normalised to a reference gene (GAPDH). The specific primers and genes for each species can be sent upon request. For all samples, TNF-α expression, or that of an equivalent gene if the sequence for the TNF-α gene was unknown for a given species, was measured after six hours incubation with LPS while the other cytokines (clinical trial) were measured after 24 hours stimulation.
Quantitative analysis was conducted on 100 ng of cDNA by qPCR using Quantifast Sybr Green® kits (Qiagen) and the ABI prism sequence detection system 7300 (Applied Biosystem) according to the manufacturer’s instructions. The PCR mixture was denatured at 95°C for 10 minutes. After this initial step, 40 amplification cycles were carried out with the following conditions for each cycle: 95°C for 15 seconds, 60°C for 30 seconds. Each PCR reaction was performed in triplicate. Results were analysed with SDS v1.2 software (Applied Biosystem) on the basis of the number of knocks of necessary genetic increase to reach the threshold of fluorescence 50% of the reference curve. Cycle threshold were transformed into relative expression “R”, which takes into account the expression of the gene of interest stimulated in vitro brought back to its native expression (without LPS stimulation), normalised against the expression of the reference gene GAPDH under the same conditions. GAPDH, a housekeeping gene is the expression of background cellular activity. Relative expression was calculated using the software program REST-MCS ©version 27 and according to the formula:
R = (E target) ΔCp target (MEAN control – MEAN sample)/ (E ref) ΔCp ref (MEAN control – MEAN sample)
In which R= Relative expression, E target = cytokine expression, E ref = GAPDH expression, ΔCp target (MEAN control – MEAN sample) = Amplification difference between control and sample for the cytokine for the sample, ΔCp ref (MEAN control – MEAN sample) = Amplification difference between control and sample for the GAPDH.
2.3 Statistics
All data were tested for normality of the distribution using the Shapiro-Wilk test, then analysed with either Mann Whitney (non parametric distribution) or Student T-test (parametric distribution) using SPSS V 17.0 Software (Paris-La Défense, France).
3. Results and discussion
In preclinical studies, we used four animal models to test the anti-inflammatory effect of Fiber 1. TNF-α expression was significantly reduced in chicken, pig and rabbit compared to the placebo group (see Figure 1A). The lack of variation in turkey may be the result of difference between activation pathways between TNF-α and IL-1β. While TNF-α is rapidly stimulated by LPS, IL-1β may take more than six hours to reach its peak of expression8. Even though the cytokines measured are not exactly the same in the different species, we noticed a significant anti-inflammatory effect of the fiber in three out of the four animal species tested. Whether the lack of effect in turkey is due to a different cytokine being measured remains to be determined, but at the time of the study IL-1β was the best candidate in the absence of a validated TNF-α sequence.
The results are consistent with the immunomodulating activity of butyrate in vitro9. We have observed that butyrate production increased in the gastrointestinal tract following those feeding regimen (data not shown). We demonstrated here that we were able to analyse the evolution of a given signal across species (see Figure 1A). If the gene and mRNA are sequenced, cytokines and inflammatory profiles can be measured in any species.
In the clinical trial, a larger panel of pro- and anti-inflammatory cytokines, effectors as well as regulatory, were measured following treatment with Fiber 2 (see Figure 1B). We observed a general trend towards normalisation of the immune signal. Fiber 2 seems to be inhibiting the LPS-induced Th1 pathway cytokines expression and normalising the LPS-induced inhibition of the Th2 pathway cytokines. The modulation however is only significant for the anti-inflammatory cytokine IL-13, even though there is a strong trend (p <0.1) for regulation of IL-1β and IL-10 as well. More cytokines were tested but no clear modulation of the cytokine expression was observed (see Figure 1B).
We can make a parallel between those observations and the immunomodulator role of probiotics10,11. It is not surprising as prebiotics can modulate bacterial populations to increase genera such as Bifidobacterium, as well as the concentration of short chain fatty acids such as butyrate (not shown). Both bifidobacteria and butyrate are widely known to be immune modulators in vitro5,12. It is interesting that this potential application for dietary fibers has only recently started to be investigated.
Tassignon et al13 presented a comparison of the different possible detection methods for cytokine modulation in a vaccination protocol. The authors showed that real time PCR quantification was more accurate than ELISA when measuring the memory response. We propose to go further and use LPS-stimulation protocol in whole blood samples rather than isolated peripheral blood mononuclear cells (PBMC). The LPS-stimulation protocol presented here is a one-step induction procedure. This efficient and rapid method enables unbiased results. Because there is no additional step to isolate a single cellular population, all blood cells and circulating elements (platelets, peptides, antioxidants) can interact in the LPS signaling cascade without barrier. Moreover and because the cells are not extracted, the use of an artificial and synthetic growth medium is not required, thus preventing any artificial background and lengthy culturing protocols14.
4. Conclusions
This protocol can be applied for zootechnic endpoints in farming as well as for investigations of immunomodulatory effects of dietary interventions to improve the quality of the meat. Indeed we are now in an economic environment where consumers aim for organic food products, free of any antibiotics and pesticides. If a dietary intervention can boost the animal’s immune system as well as improve the meat output, there will be an increasing demand in scientific proof to back up such allegations.
In a clinical aspect, we can assess the dietary impact of a large number of nutritional factors, including fibers, and investigate the mechanisms of action of such modulators. As the early years of life are crucial for the development of the immune system, this potential role of fibers is comforting the current use of oligosaccharides in infant formulas. Alternatively, there is also a potential in reducing or preventing the low grade inflammatory damages associated with our lifestyles, such as smoking, low antioxidant intake and so on, as well as a therapeutic potential to relief symptoms associated with acute or chronic inflammation states, such as in diabetes, obesity, inflammatory bowel disorders or allergies.
The methodology for ex-vivo stimulation of whole blood associated with PCR detection of gene expression is a tool in the search to better understand those health effects. It offers benefits against classical ex-vivo stimulation protocols by being a one-step process, thus avoiding the drawbacks of leukocyte isolation. The isolation of culturing of leukocytes in a standardised culture medium does not allow for the possible impact of blood environment and other blood cells in the signaling process induced by LPS stimulation.
References
- Bjorksten B. Disease outcomes as a consequence of environmental influences on the development of the immune system. Curr Opin Allergy Clin Immunol 2009;9:185-9.
- Calder Pc, Krauss-Etschmann S, De Jong Ec, Dupont C, Frick Js, Frokiaer H, Heinrich J, Garn H, Koletzko S, Lack G, Mattelio G, Renz H, Sangild Pt, Schrezenmeir J, Stulnig Tm, Thymann T, Wold Ae and Koletzko B. Early nutrition and immunity – progress and perspectives. Br J Nutr 2006;96:774-90.
- Cashman Kd and Shanahan F. Is nutrition an aetiological factor for inflammatory bowel disease? Eur J Gastroenterol Hepatol 2003;15:607-13.
- Dutau G and Rancé F. The story of food allergy from its first recognition to the present time. Rev Fr Allergol Immunol Clin 2006;46:312-23.
- Dossus L, Becker S, Achaintre D, Kaaks R and Rinaldi S. Validity of multiplex-based assays for cytokine measurements in serum and plasma from “non-diseased” subjects: comparison with ELISA. J Immunol Methods 2009;350:125-32.
- Neuder Le, Keener Jm, Eckert Re, Trujillo Jc and Jones Sl. Role of p38 MAPK in LPS induced pro-inflammatory cytokine and chemokine gene expression in equine leukocytes. Vet Immunol Immunopathol 2009;129:192-9.
- Pfaffl Mw, Horgan Gw and Dempfle L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res 2002;30:e36.
- Bird S, Zou J, Wang T, Munday B, Cunningham C and Secombes Cj. Evolution of interleukin-1beta. Cytokine Growth Factor Rev 2002;13:483-502.
- Siwicki Ak, Fuller Jc, Jr., Nissen S, Ostaszewski P and Studnicka M. In vitro effects of beta-hydroxy-beta-methylbutyrate (HMB) on cell-mediated immunity in fish. Vet Immunol Immunopathol 2000;76:191-7.
- Dugas B, Mercenier A, Lenoir-Wijnkoop I, Arnaud C, Dugas N and Postaire E. Immunity and probiotics. Immunol Today 1999;20:387-90.
- Rautava S, Kalliomaki M and Isolauri E. New therapeutic strategy for combating the increasing burden of allergic disease: Probiotics-A Nutrition, Allergy, Mucosal Immunology and Intestinal Microbiota (NAMI) Research Group report. J Allergy Clin Immunol 2005;116:31-7.
- Van Hoffen E, Korthagen Nm, De Kivit S, Schouten B, Bardoel B, Duivelshof A, Knol J, Garssen J and Willemsen Le. Exposure of Intestinal Epithelial Cells to UV-Killed Lactobacillus GG but Not Bifidobacterium breve Enhances the Effector Immune Response in vitro. Int Arch Allergy Immunol 2009;152:159-68.
- Tassignon J, Burny W, Dahmani S, Zhou L, Stordeur P, Byl B and De Groote D. Monitoring of cellular responses after vaccination against tetanus toxoid: comparison of the measurement of IFN-gamma production by ELISA, ELISPOT, flow cytometry and real-time PCR. J Immunol Methods 2005;305:188-98.
- Ocmant A, Michils A, Schandene L, Peignois Y, Goldman M and Stordeur P. IL-4 and IL-13 mRNA real-time PCR quantification on whole blood to assess allergic response. Cytokine 2005;31:375-81.
About the author
Afif Abdel Nour has a PhD in Nutrigenomics and specialises in applied techniques in Molecular Biotechnology. He did his Masters in Molecular Microbiology from Pasteur Institute Lille. Currently, he is working as a Biotechnology lecturer and Molecular Biotechnology consultant at Institute Polytechnique LaSalle Beauvais, France and in several countries in the Middle East.
